
Pedro Diaz-Vivancos, Associate Researcher (CEBAS-CSIC)
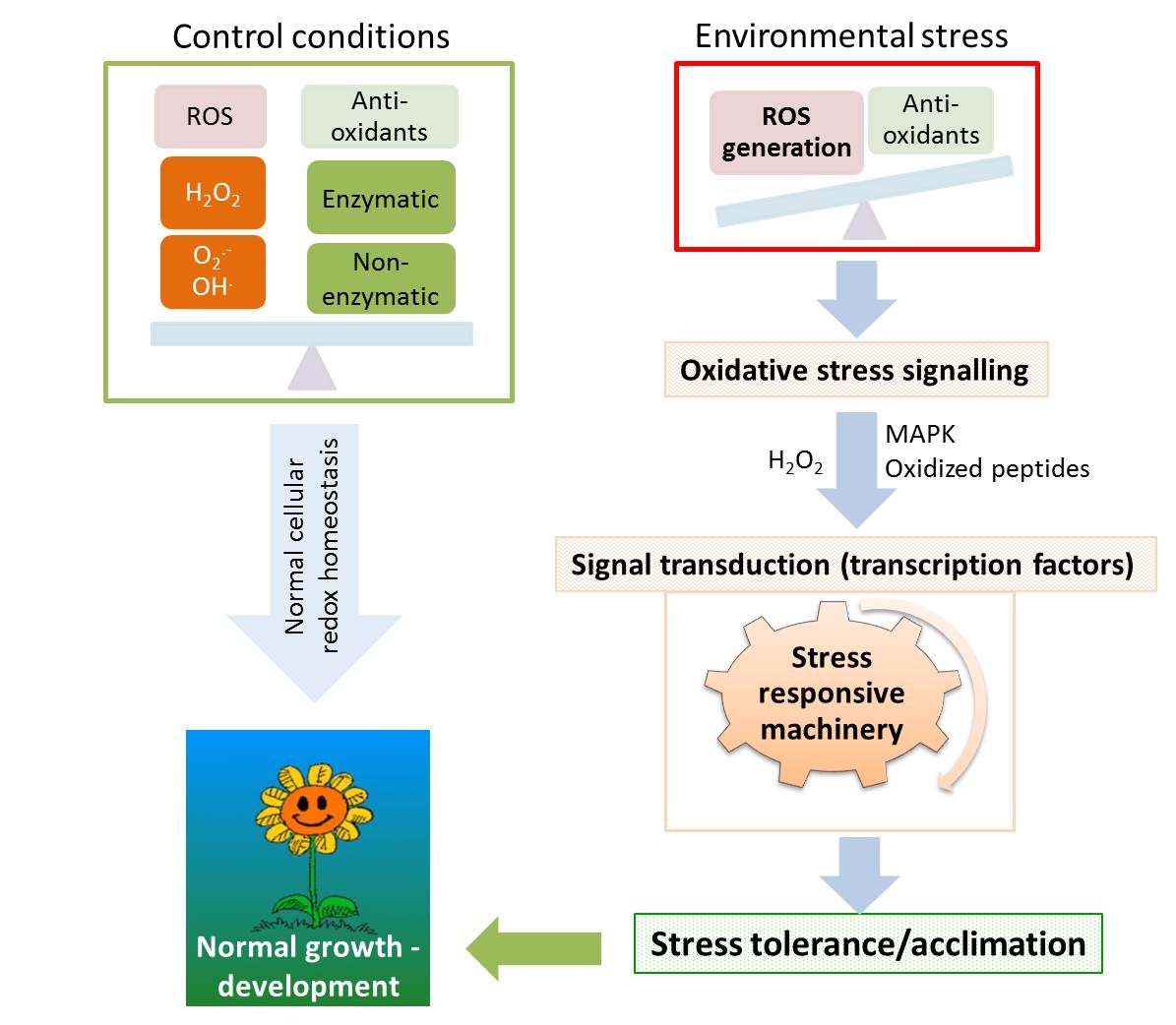
Plants are sessile forms of life and as consequence they don´t have the capacity to “escape” from unfavourable environmental situations. Thus plants often face the challenge of grow under stressful conditions such as water or light deficit or excess, low or high temperature, pathogen attack… These stresses exert adverse effects on plant growth and development by inducing many metabolic changes, such as the occurrence of an oxidative stress (Diaz-Vivancos et al., 2008; Hernández et al., 2004a,b). Oxidative stress involves the enhanced production of reactive oxygen species (ROS), such as superoxide (O2•-), hydrogen peroxide (H2O2), and hydroxyl radicals (OH.), which are some of the most damaging molecules in plants.
In plants, ROS production is kept under tight control by an efficient antioxidative system, which includes both enzymatic and non-enzymatic compounds, that modulates intracellular ROS concentration setting the cellular redox homeostasis. Among these non-enzymatic scavengers, low molecular weight compounds, including ascorbic acid (ASC) and glutathione (GSH) are involved, while the main enzymatic arsenal of ROS scavengers includes enzymes such as superoxide dismutase (SOD), catalase (CAT), peroxidase (POX) and the ascorbate-glutathione cycle enzymes [ascorbate peroxidase (APX), dehydroascorbate reductase (DHAR), monodehydroascorbate reductase (MDHAR) and glutathione reductase (GR)]. Under environmental stress conditions antioxidants function as redox buffers that interact with ROS and act as a metabolic interface that modulate the appropriate induction of acclimation/tolerance responses (Foyer & Noctor 2005).
Acclimation of plants to changes in their environment requires a new state of cellular homeostasis achieved by a delicate balance between multiple pathways that reside in different cellular compartments. Under optimal growth conditions, ROS are mainly produced at a low level, being their rate of production drastically increased when plants are exposed to environmental stresses. Despite the fact that ROS has been considered as toxic molecules during stress conditions, recent studies have shown that ROS play a key role in plants as signal transduction molecules acting as signalling molecules that regulate stress responses, as well as growth and development (Foyer & Noctor 2005; Miller et al., 2010; Barba-Espín et al., 2011).
The next question is obvious, which ROS is the most suitable to be messenger? H2O2 is the most likely ROS to act as messenger because of its relative stability and its capacity to cross membranes, thus H2O2 is considered to play a major role in cellular signalling pathways (Foyer & Noctor 2005). Additionally, H2O2 lead to the activation of mitogen activated protein kinases (MAPKs; a specific class of serine/threonine protein kinases) cascades which play a key role in plant growth and development (Barba-Espín et al., 2011) as well as in biotic and abiotic responses. Moreover, oxidized peptides could act as secondary ROS messengers regulating source specifics genes and might contribute to ROS signalling during oxidative stress (Møller & Sweetlove 2010).
The oxidative stress that accompanies environmental stresses appears now to be a key component in plant cell signalling. Therefore, the oxidative stress should not necessarily be viewed as a harmful event needed to be avoided or alleviated, but could also be viewed as a perquisite for the plant to adequately respond and induce proper acclimation mechanisms. Because it seems that ROS signalling is responsible for the activation of different acclimation responses in plants, it is easy to understand why changes in ROS metabolism were found to cause enhanced tolerance or sensitivity to stress situation (Miller et al., 2010).
References:
Barba-Espin G et al. (2011) Understanding the role of H2O2 during pea seed germination: a combined proteomic and hormone profiling approach. Plant, Cell and Environment 34: 1907-1919.
Diaz-Vivancos P et al. (2008) Alteration in the chloroplastic metabolism leads to ROS accumulation in pea plants in response to Plum pox virus. Journal of Experimental Botany 59: 2147-2160.
Foyer CH & Noctor G (2005) Redox homeostasis and antioxidant signalling: a metabolic interface between stress perception and physiological responses. The Plant Cell 17: 1866-1876.
Hernández JA et al. (2004a) Oxidative stress induced by long-term plum pox virus infection in peach (Prunus persica L. cv GF305). Physiologia Plantarum 122: 486-495
Hernández JA et al. (2004b) Role of hydrogen peroxide and the redox state of ascorbate in the induction of antioxidant enzymes in pea leaves under excess light stress. Functional Plant Biology 31: 359-368.
Miller G et al. (2010) Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant, Cell and Environment 33: 453-497.
Møller IM & Sweetlove LJ (2010) ROS signalling – specificity is required. Trends in Plant Science 15: 370-374.
